1. 서 론
한국 산림의 주요 수종인 소나무는 과거부터 가장 선호되는 건축 자재이었으며, 우리나라 목조건축문화재에는 대부분 소나무가 사용되어왔다(Park and Lee, 2007). 따라서 우리나라 문화재 수리 표준시방서에는 “문화재수리에 사용하는 목재는 기존 부재를 최대한 재사용하되, 부식이 심하거나 손상되어 재사용이 불가한 것은 육송 또는 동등 이상의 품질을 가진 목재를 사용한다.”라고 명시되어 있다. 현재 목조건축문화재 수리 시 교체가 필요한 부재에 한해 대부분 국내산 소나무(Pinus densiflora S. et Z.)로 교체되고 있으나, 일부 외래산 소나무가 혼입되어 사용된 경우가 확인되었다(Cultural Heritage Conservation Science Center, 2016). 이는 국내산과 일부 외래산 소나무의 해부학적 구조가 유사하기 때문에 교체부재에 대한 식별 검증이 어려움으로 인해 발생된 것이다. 따라서 국내산 소나무와 해부학적 구조에 차이가 없는 외래산 소나무(Pinus resinosa, Pinus sylvestris)를 식별할 수 있는 연구가 필요하다.
과거 외래산 소나무 식별은 대부분 해부학적 구조 관찰 방법으로 접근하였으나 국내산 소나무와 유사한 해부학적 구조로 인해 한계가 있었다. 따라서 DNA를 이용한 방법, 근적외선을 이용한 방법 및 영상장치를 이용한 방법과 같이 다양한 방법들이 현재 연구되고 있으며, 특히 DNA를 이용한 방법의 경우 외래산 소나무 식별 가능성은 확인되었지만 정확히 식별하여 연구결과가 도출된 사례는 현재 확인되지 않고 있다(Hwang, 2015; Yoo et al., 2020; Hong et al., 2014; Kim and Jung, 2007; Yang, 2019). 따라서 외래산 소나무를 식별할 수 있는 다른 방법으로의 접근이 필요하다.
소나무를 구성하고 있는 세포 중 가도관은 90% 이상을 차지하고 있으며, 2-4 mm 길이를 가지고 있다(Park et al., 1987). 또한 이전 연구에서 ‘소나무를 구성하고 있는 가도관 길이의 변화는 임목이 갖는 생물학적 특징에 의해 결정된다’라고 하였고(Seo et al., 2014), ‘수목이 생장하고 있는 기후조건에 의해 세포의 마이크로피브릴 각도를 비롯한 해부학적 특성이 지역적으로 차이가 존재한다’라고 하였다(Kim et al., 2020). 따라서 국내산 소나무와 유사한 해부학적 구조를 가지는 외래산 소나무는 생물학적 특징의 차이뿐만 아니라 다른 기후 및 환경 조건에서 생장하였으므로 연륜폭과 연륜을 구성하는 세포 크기의 차이도 있을 것으로 판단하였다.
따라서 본 연구에서는 국내산 소나무와 유사한 해부학적 구조를 가지는 외래산 소나무를 대상으로 연륜해부학적 방법으로 식별하고자 국내 지역별 연륜연대기를 구축하고 가도관 및 단열방사조직 세포의 크기를 측정 및 비교를 통해 외래산 소나무를 식별할 수 있는 가능성을 확인하고자 하였다.
2. 재료 및 방법
2.1. 조사지 및 시료채취
국내산 소나무 시료는 우리나라 주요 국립공원 13곳에 자생하는 수고 7-20 m, 흉고직경 39-72 cm 이상의 성숙재 20본을 채취하였다. 각 국립공원에서 채취된 20본의 시료 중 연륜개수가 50개 이하인 시료를 제외한 11-20본의 시료가 연륜분석에 사용되었다(Table 1). 시료는 이상재를 피할 수 있는 흉고의 양방향에서 생장추를 이용하여 코어시료 형태로 채취하였다. 국내산 소나무와 유사한 해부학적 구조를 가지는 외래산 소나무 시료들을 확보하고자 전국에 목재 수입 및 가공 사업장들을 대상으로 진행하였으나, 국내산 소나무와 해부학적으로 분류가 되는 Ponderosa group (Pinus radiata류)과 Taeda group (Pinus rigida류)들이 대부분이었다. 또한 2008년 제정된 식물방역법으로 인해 수입되는 소나무들은 대부분 원목이 아닌 가공된 형태로 약제 및 열처리가 진행되어 수입되고 있기 때문에 정확한 벌채연도를 확인하기 어렵고, 연륜연대학적 분석을 위한 충분한 연륜수를 가지고 있는 시료 확보가 매우 어려웠다. 따라서 전국의 사업장 중 Sylvestris group 소나무를 수입 및 가공하는 업체 2곳만이 확인되어 러시아와 유럽에서 벌채되어 수입된 것으로 추정되는 시료를 디스크의 형태로 각각 30점씩 채취할 수 있었다(Table 1).
2.2. 분석방법
2.2.1. 수종분석
러시아와 유럽에서 벌채되어 수입된 것으로 추정되는 소나무 시편 각 30점씩 총 60점에 대해 모두 수종분석을 진행하였다. 면도날을 이용하여 3단면 박편을 제작하여 수종 분석용 프레파라트를 제작한 후 광학현미경(ECLIPSE LV100, Nikon, JPN)으로 관찰하였다.
2.2.2. 연륜폭 측정
국내산 소나무 시료들은 코어 형태로 채취하였으므로 연륜 고정대에 부착하였고, 외래산 소나무는 디스크 형태로 별도의 전처리 없이 진행하였다. 각 시료는 밸트샌더로 연마하여 연륜의 경계가 선명하게 관찰될 수 있도록 하였다. 각 연륜의 폭은 컴퓨터에 부착된 연륜폭측정기(Lintab, Rinntech, DEU)를 이용하여 0.01 mm 정확도로 측정하였다. 정확한 생육연대를 각 연륜에 부여하기 위해 크로스데이팅을 실시하였다. 크로스데이팅은 연륜연대학에서 자주 사용되는 TSAP-Win 프로그램을 사용하였고, 생장경향을 비교하는 그래프방법(graphic method)과 상관계수를 이용하여 계산된 t값 및 부호검정을 응용한 G값(gleichläaufigkeit) 등의 통계 기능을 이용하였다(Rinn, 2011; Schweingruber, 1988; Baillie and Pilcher, 1973; Eckstein and Bauch, 1969). 100년 이상의 연대기를 상호 비교할 때, 유의성 있는 결과의 기준은 t값 3.5 이상이며, G값 65% 이상이다(Park et al., 2003).
2.2.3. 연륜별 시료 채취 및 세포 측정
크로스테이팅을 통해 국내의 13개 지역별 소나무 연대기와 2개의 외래산 소나무연대기가 작성되었다. 각 연대기에 포함된 시료들 중 통계적 유의성이 가장 높은 시료 3점씩을 선별하였다. 이전 연구에서 가도관 길이가 안정화되는 기간은 40년이었다(Seo et al., 2014). 따라서 선별된 시료를 관찰하여 수에서부터 40년 이상이 지난 변재부위에서 30년(1987-2016년)간 연륜을 조재와 만재로 구분하여 면도날로 재단하였다(Figure 1A). 재단된 시료들의 가도관을 측정하기 위해 Schurz 용액(질산 35% 100 ml + 염소산칼륨 6 g)에 일주일간 담가두어 해리하였다. 해리된 가도관들은 증류수를 이용하여 3회 세척하였으며, 정확한 가도관 길이 측정을 돕기 위하여 샤프라닌으로 염색하고 프레파라트를 제작하였다(Figure 1B). 단열방사조직의 높이와 각 단열방사조직당 구성 세포수 측정을 위해 방사방향 폭을 고려하여 3년 간격으로 1987년부터 2016년까지 총 10개의 시료를 제작하였고, 시료의 접선단면에서 박편을 채취하여 프레파라트를 제작하였다.
제작된 각 프레파라트에서 가도관과 단열방사조직을 30개씩 선택한 후 광학현미경에 부착된 이미지 분석프로그램을 이용하여 0.01 mm단위로 측정하였고, 각 단열방사조직당 구성 세포수도 측정하였다(Figure 1C).
3. 결과 및 고찰
3.2. 연륜연대분석
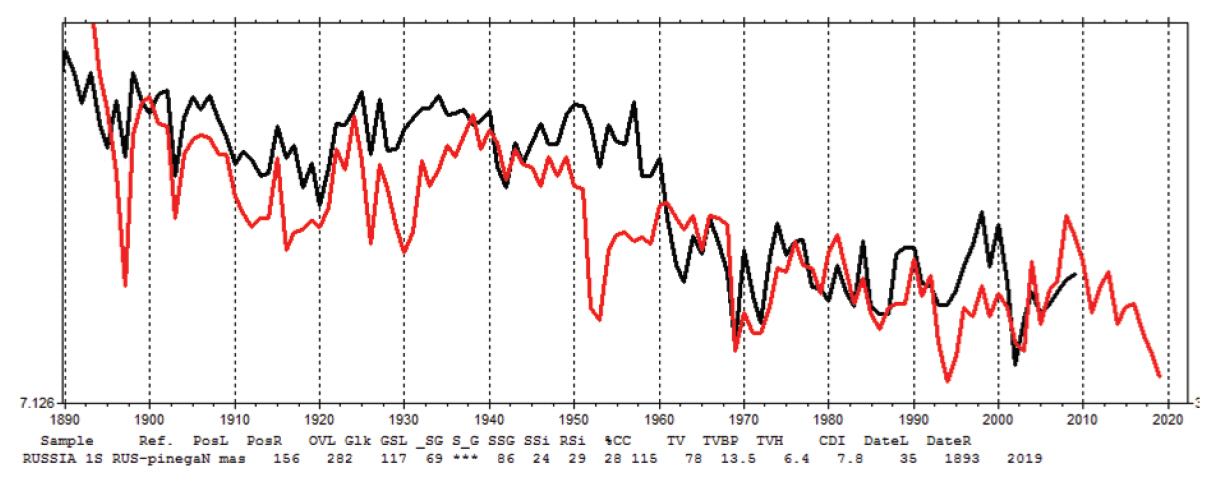
러시아와 유럽에서 벌채되어 수입된 것으로 추정되는 각 30점의 소나무 연륜패턴은 높은 통계적 유의성을 보였으며, 각각 119년, 112년의 소나무연륜연대기를 작성할 수 있었다. 작성된 연륜연대기를 이용하여 벌채지역과 벌채시기를 확인하고자 우선 국내 소나무 표준연륜연대기와 크로스데이팅을 실시한 결과, 통계적 유의성이 낮고 그래프도 일치하지 않는 것으로 확인되었다. 따라서 국내 소나무와 유사한 특징을 가지는 P. sylvestris, P. resinosa, P. kesiya, P. merkusii, P. nigra의 28개국 396개 표준 연륜연대기를 NOAA(National Oceanic and Atmospheric Administration)에서 제공받아 크로스데이팅하였다. 러시아에서 수입된 것으로 추정되는 소나무 연륜연대기는 러시아 Verhnaja Toima와 Arkhangelsk 지역의 P. sylvestris 연대기와 t값 4.9, G값 69% 이상으로 비교적 높은 통계적 유의성을 나타내었으며, 육안상으로도 그래프가 서로 일치하는 것이 확인되었다(Figure 3). 또한 유럽 소나무 연륜연대기는 독일 Hessen 지역과 Nordrhein-Westfalen 지역 연대기와 t값 5.1, G값 64% 이상으로 비교적 높은 통계값을 나타내었으며, 육안상으로도 그래프가 서로 일치하는것이 확인되었다(Figure 4). 따라서 러시아와 유럽에서 벌채되어 수입된 것으로 추정된 외래산 소나무들은 러시아 모스크바 북부지역과 독일 중서부지역 주변에서 자생하고 있었으며, 2019년에 벌채되어 가공 후 2020년과 2021년에 각각 수입된 것으로 판단할 수 있었다.
3.3. 가도관 길이
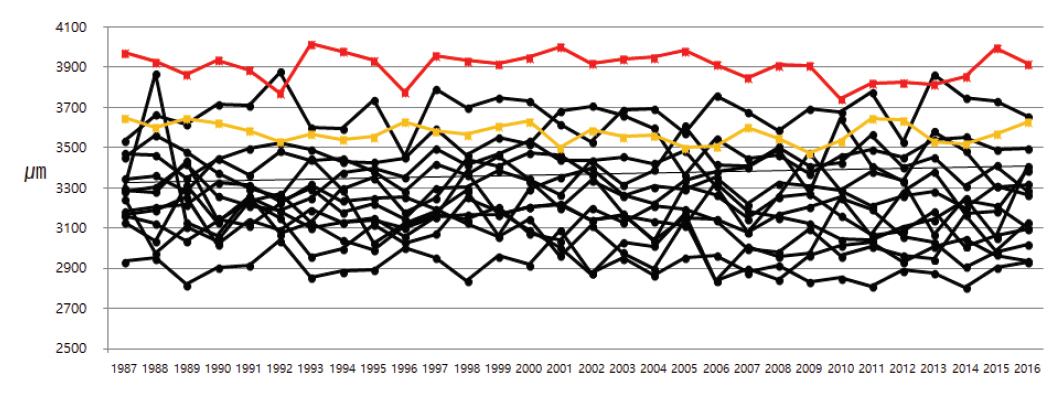
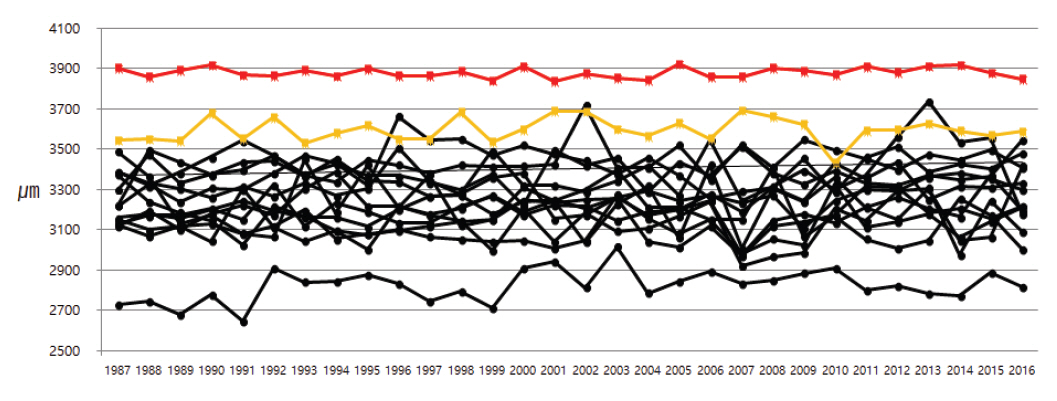
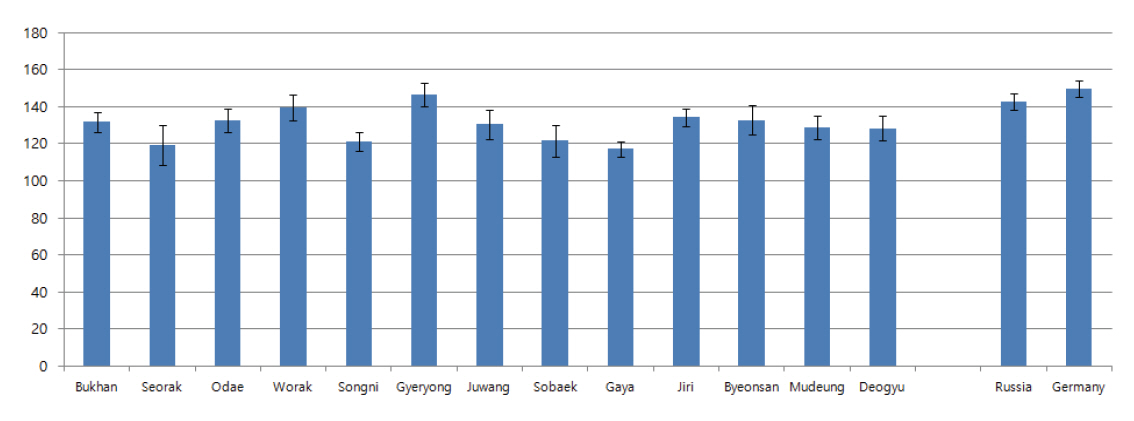
국내 각 국립공원에서 자생하는 소나무들의 30년간 연륜별 가도관 길이는 매년 차이는 있지만 성숙재에서 채취하였으므로 길이가 증가하는 변화는 관찰되지 않았다. 국내 소나무의 연륜별 조재 가도관 평균 길이는 2.74-3.88 mm 사이로 측정되었고, 만재 가도관 평균 길이는 2.64-3.73 mm 사이로 측정되었다. 반면 러시아 소나무의 연륜별 조재 가도관 평균 길이는 3.74-4.02 mm 사이로 측정되었고, 만재 가도관 평균 길이는 3.84-3.92 mm 사이로 측정되었으며, 유럽 소나무의 조재와 만재 가도관 평균 길이는 3.43-3.69 mm와 3.47-3.65 mm 사이로 측정되었다(Figure 5, 6). 국내 소나무의 가도관 길이를 측정한 이전 연구들에서도 성숙재의 가도관 길이는 조재의 경우 3.25-4.07 mm, 만재는 3.15-3.81 mm으로 측정되어 본 연구에서 측정된 결과와 거의 일치하였다(Seo, et al., 2014; Kim et al., 2018; Kim and Kim, 2018). 또한 폴란드, 라트비아, 핀란드에서 자생하는 P. sylvestris 성숙재 가도관 길이를 연구한 이전 연구들에서도 국내 소나무 가도관 길이와 유사하게 약 2.7-3.8 mm 사이로 측정되었다(Irbe et al., 2013; Mäkinen and Hynynen, 2014; Fabisiak and Fabisiak, 2021). 하지만 본 연구에서는 러시아 소나무의 가도관 길이가 표준편차 내에서 벗어나 차이가 있는 것으로 확인되었다(Figure 7). 러시아에서 수입된 P. sylvestris의 변재부 가도관 길이를 측정한 이전 연구에서도 평균 길이가 4.00 mm로 본 연구 결과와 유사한 결과가 확인되었고, 북양재의 경우 전체적으로 국내 수종들에 비해 가도관 길이가 약간 긴 경향이 있는 것으로 평가하였다(Lee, 1990). 따라서 추가연구가 필요하겠지만 외래산 소나무 중 일부 소나무들은 국내산과 가도관 길이 차이를 보이며, 이는 당시 소나무가 자생하고 있는 지역적인 특징(기후, 토양, 주변식생 등)에 의한 것으로 판단된다.
3.4. 단열방사조직 높이
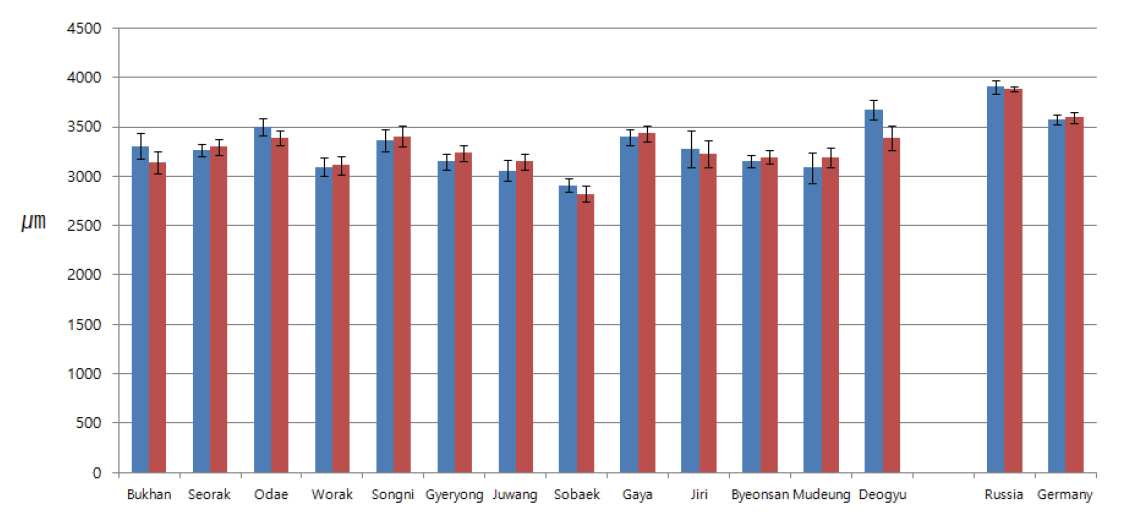
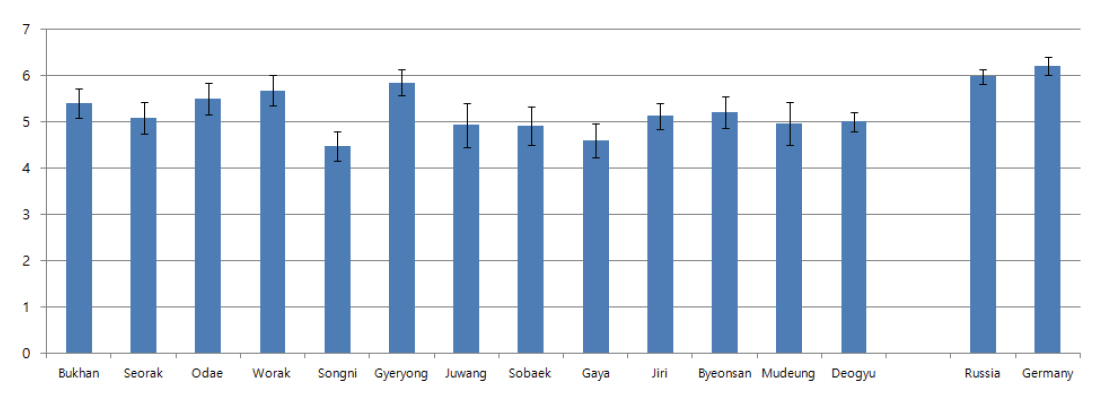
접선단면상에서 국내 소나무들의 단열방사조직 높이는 대부분 120-140 μm이었고, 축방향을 기준으로 각 단열 방사조직당 4.4-5.5개의 세포로 구성되어 있었다(Figure 8, 9). 국내 소나무의 단열방사조직의 높이를 측정한 이전 연구에서는 이상재가 있는 부위에서 약 155-170 μm 사이로 확인되었고(Chung and Lee, 1989), 국내 소나무와 금강송에 대한 각 단열방사조직당 구성세포수를 측정한 연구에서는 5.7-6.3개로 측정되었다(Kim et al., 2014). 본 연구 결과가 이전 연구들에 비해 다소 낮게 측정되었는데, 이는 이전 연구들이 압축이상재 또는 수부분과 심재부를 포함한 부분에서 측정된 결과이기 때문에 변재부위에서만 측정한 본 연구와 차이가 있는 것으로 판단된다.
러시아와 유럽에서 벌채된 것으로 추정되는 외래산 소나무의 방사조직 높이는 143 μm, 각 단열방사조직당 구성세포수는 6.0개 이상을 나타내어 국내 월악산과 계룡산 소나무와 유사한 경향을 나타내었다. 반면에 나머지 지역의 국내 소나무들보다는 상대적으로 단열방사조직의 높이가 높고 각 단열방사조직당 구성세포 수가 많은 경향을 나타내었다. 그러나 표준편차를 고려했을 때 국내산 소나무와 외래산 소나무의 단열방사조직 크기 차이는 없는 것으로 판단되었다.
4. 결 론
국립공원 13곳에서 자생하는 국내산 소나무와 유사한 해부학적 구조를 가지는 외래산 소나무의 연륜 및 세포크기분석을 통해 연륜해부학적 방법으로 외래산 소나무를 식별하고자 하였다. 국내와 기후 및 환경이 다른 지역에서 자란 외래산 소나무의 연륜연대기는 국내 소나무 표준연륜연대기와 일치하지 않았고, 외국의 소나무 표준연륜연대기와 높은 통계적 유의성을 보여 벌채된 국가 및 인접지역, 정확한 벌채시기까지 확인할 수 있었다. 국내산과 외래산 소나무의 가도관과 단열방사조직 높이를 비교한 결과에서는 러시아에서 벌채되어 수입된 소나무들의 가도관 길이를 제외하고는 차이가 없는 것으로 확인되었다.
현재 가도관과 단열방사조직의 크기에 대한 연구는 진행중에 있으며, 연구가 완료되는 즉시 국내산과 외래산 소나무의 차이뿐만 아니라 국내 지역별 차이를 통계적으로 분석 및 분류하여, 현재보다 더 정확한 결과를 제시하고자 한다.